botanik
-
İçerik sayısı
297 -
Katılım
-
Son ziyaret
-
Bitkiler toplanıp, kurutulduktan sonra (bu safhaların da ayrıntılı teknikleri vardır) herbaryum kartonlarına yapıştırılır ve sıra bunların adlandırılmasına gelir. Bitki örnekler şu kaynaklardan yararlanılarak adlandırılabilirler: 1- Floralar: Türkiye için E.Bossier’in “Flora Orientalis” ve P.H.Davis’in “Flora of Turkey” adlı eserleri temel kaynaktır. 2- Monograf ve Revizyonlar: Bir taksonun (familya, genus gibi) tüm yeryüzü türlerini inceleyen taksonomik araştırmaya monografi, belirli bir alanın türlerini inceleyene de revizyon denir. Revizyonlar genellikle Floralara hazırlık için yapılır. 3- Herbaryumlar 4- Resim ve Şekiller: Bitkilerin resim ve şekillerini içeren kitap ve dergilerden yararlanılır. Bu kaynaklardan yalnız birinden yararlanılarak yapılan adlandırılmaların problemleri vardır. O nedenle en doğru adlandırma metodu Flora veya revizyon kullanılarak yapılan adlandırmalardan sonra herbaryumlardan da kontrol etmektir. Herbaryumlara bakılırken eski örneklerin kim tarafından adlandırıldığına dikkat edilmelidir. Resim ve şekillerden adlandırma çok kullanılan bir metot olmamakla birlikte tanıma yardımcı bilimsel şekiller ve resimler bulununca özellikle çok karakteristik ve gösterişli çiçekli bitkiler için kullanılabilir. Flora, revizyon ve monografilerden bitki adlandırılırken, tayin anahtarından yararlanılır. Tayin anahtarları bitki türleri veya taksonları arasındaki farklı morfolojik karakterlerden yararlanılarak hazırlanır. Anahtarlar temel olarak ikili-çatal (dikotomik) tiptedir. Bazen bunun yanında formüller (çok girişli, multi-acsess) anahtar da kullanılmaktadır. İkili-çatal anahtar hazırlanırken aynı karakterin farklı şekillerinden faydalanılır. Yani her durum için bir karakter ile bunların zıt özellikleri kullanılır (örneğin; petaller sarı – petaller kırmızı. Burada kullanılan karakter petal rengidir, farklı yönleri ise petalin sarı veya kırmızı olmasıdır).Türkiye Florasında kullanılan anahtar Dikotom=Çatallı Anahtardır. Ancak Umbelliferae, Compositae, Graminae familyaları için Formül (Multi-Access= Çok Girişli) anahtarlar da hazırlanmıştır. Yurdumuz bitkileri üzerinde yapılan yaygın arazi çalışmaları uzun yıllar Tohumlu Bitkiler ile Eğreltiler üzerinde yoğunlaştırılmıştır. Yosun, Mantar ve Likenler üzerindeki çalışmalar ise son yıllarda yerli botanikçiler tarafından yoğunlaştırılmış ise de bu gruplar üzerindeki çalışmalar Tohumlu Bitkilere göre henüz oldukça azdır. Algler üzerinde de son yıllarda, özellikle son 20 yıl içinde oldukça yaygın çalışmalar yapılmış, hem deniz hem de tatlı su ekosistemleri incelenmeye başlanmıştır.
-
Her türün insanlarda olduğu gibi bir adı vardır ve bu ad 2 kelimeden oluşur. Bunlardan birincisi o canlının cins adı, ikincisi ise tür adıdır ve bu adlar sistematiğin başlangıç yılı olan 1753’den beri latince olarak ve iki isimli (binominal ) yazılır. Örneğin dağlarımızda doğal yetişen pembe gül’ün adı Rosa canina’dır. Burada Rosa gül cinsinin, canina ise tür’ün adı olup tür adıdır. Esasında bu adın tam yazılışı bu isimlerin sonuna, bu türü dünyaya ilk tanıtan kişinin adı veya kısaltılmışı da eklenerek yazılması gerekir. Örneğin pembe gül’ün adının tam yazılışı Rosa canina L. Olmalıdır. L. Bu türü tanıtan kişinin yani Linneaus (Linne okunur)’un adının kısaltılmış şeklidir. Herhangi bir yazı içinde bitkilerin latince isimleri, dikkat çekmek için, italik harflerle yazılır. Bitkiler 1700 lü yıllara kadar kısa tanımları sayılabilecek birkaç kelime ile adlandırılırdı. Örneğin yumurtamsı yapraklarının kenarları sivri dişli kavak gibi. Linne’nin Species Plantarum adlı eserinin yayın tarihi olan 1753 den beri bitkiler iki adla anılmakta ve bu ad bütün dünyada aynı olmaktadır. Bunlardan birincisi cins adıdır, ikincisi epitet, yani tür adıdır. Bu sisteme binominal sistem denir. Örnek: Pyrus malus L. Elma Bitkinin cins adı daima büyük harfle, tür adı (epiteti) ise daima küçük harfle başlar. Bitki adlarının sonuna, bilim dünyasına ilk tanıtan kişinin adı veya kısaltılmışı yazılır. Adı tek harfle kısaltılan tek kişi Linne’dir. Epitet, iki isimli olması durumunda Capsella bursa-pastoris (L.) Medik şeklinde yazılarak son iki isim birleştirilir. Türün, eğer varsa tür altı kategori yani alttür (subspecies) ve varyete adları da daima küçük harfle başlar. Örnek: Juniperus communis L. subsp. nana Syme (Cüce ardıç) Bitkilere verilen tür adları (epitet’ler), bitkinin ya morfolojik özelliklerinden, ya yetiştiği coğrafik veya ekolojik ortamlardan, ya bir kişinin ya da yer adından gelir. İsimlerin latince olma zorunluluğu olduğundan verilen isimler latinceleştirilir. Juniperus foetidissima Willd. (Kokar ardıç) Juniperus drupacea Lab. (meyvesi drupa tipinde) Juniperus phoenicea L. (Finike ardıcı) Ferula longipenduculata Peşmen (uzun pedunkullu Çakşır) Ferula halophila Peşmen (tuzcul alanda yetişen Çakşır) Iris nectarifera A.Güner (Nektar taşıyan süsen) Abies nordmanniana (Stev.) Spach (Nordmann Göknar’ı) Pentapera bocquetii Peşmen (Bocquet’den) Geranium davisianum Peşmen et Güner (Davis’den) Potentilla anatolica Peşmen (Anadolu’dan) Lathyrus bitlisicus Peşmen (Bitlis Burçağı) Aubrieta anamasica Peşmen et Güner (Anamas’dan) Yetişme ortamından türetilmiş epitetler: Pinus sylvestris L. (sarıçam, ormandan) Ferula halophila Peşmen (Çakşır, tuzculdan) Kişi adından türetilmiş epitetler: Abies nordmanniana (Stev.) Spach Pentapera bocquetii Peşmen et Güner Ekimia bornmuelleriana (hem cins, hem de tür adı 2 ayrı botanikçiye izafeten verilmiştir. Coğrafik yer adından türetilmiş epitetler: Juniperus phoenicea L. (Finike’den) Potentilla anatolica Peşmen (Anadolu’dan) Lathyrus bitlicus Peşmen (Bitlis’ten) Aubrieta anamasica Peşmen et Güner (Anamas’dan) Coğrafik yön adından türetilmiş epitetler: Picea orientalis (L.) Link (doğulu) Linnea borealis L. (kuzeyli’den) Celtis autralis L. (güneyli’den) Helianthemum occidentale Willk. (batılı) Tip Örnek: Bir bitki örneğinin dünyaya tanıtılması sırasında kullanılan ve genellikle o bitkiyi en iyi şekilde temsil ettiğine inanılan örnektir. O bitkini toplandığı yere tip lokalite veya Locus Typicus denir. Aynı bitki birden fazla kişi tarafından farklı adlarla tanımlanırsa, bunlardan ilk tanım geçerli kabul edilir, öbürleri eş ad (sinonim) olur. Örnek: Pinus nigra Arn. (1785), Sin.: Pinus fenzlii Ant. et Kotschy (1867) ÇOK ÖNEMLİ: Bir yazı içinde geçen latince bitki isimleri, diğer yazılardan kolay ayrılması için, diğer yazılardan farklı (italik veya koyu vb.) yazılır.
-
Sınıflandırma tartışılmaz bir ihtiyaçtır. Bu ihtiyaç en azından canlıları adlandırma, sonra onları tanıdık kategoriler halinde gruplandırma gereksiniminden kaynaklanır. Ancak böylece mevcut çeşitliliği araştırabilmeye ve anlamaya başlayabiliriz. Bu ihtiyacı sadece taksonomistler ve biyologlar değil, normal insanlarda duyarlar. Zaten bu yüzden sınıflandırma, insanoğlu ile birlikte içgüdüsel olarak başlamıştır. Sınıflandırma eyleminde, birbirine benzer bireyler bir gruba toplanır. Daha sonra bu gruplar kendi aralarındaki benzerliklere göre daha büyük gruplar halinde bir araya getirilir. Bu işlem gittikçe büyüyen ve genişleyen gruplar halinde, tüm canlılar tek bir grup halinde toplanana kadar yükselerek sürer. İşte bu tip sınıflandırmalara Hiyerarşik Sınıflandırma denir. Bir hiyerarşideki düzeylerin, teorik olarak bir sınırı yoktur. Uluslararası Botanik Adlandırma Koduna göre temel kategoriler (düzeyler) aşağıdan yukarı doğru şunlardır: Tür, Cins, Aile, Takım, Sınıf, Bölüm, Alem. Bu kategoriler arasında başka kategoriler ve bunların ast ve üst birimleri bulunabilir. Botanikte kullanılan tüm kategoriler ve varsa düzeylerini belirleyen sonekleri ve birer örnek isim aşağıda verilmiştir.
-
Sınıflandırma sistemleri tarihinin, bilimin tarihi kadar eski olduğunu söylemek yanlış olmaz. İlk insan deneme yanılma yolu ile canlıları kendine yararlı ve zararlı olanlar diye sınıflandırmış olmalıdır. Sistematiğin yani sınıflandırmanın ilk ortaya çıkışından bugüne kadar pek çok sistemler ortaya çıkmış olmakla birlikte bu sistemler aşağıdaki gibi gruplara ayrılabilir: Yapay (Ampirik) Sistemler (M.Ö. 3000 – M.S.1580) veya Bitkilerin dış görünüşüne (HABİTUS’a) dayanan sınıflandırmalar: Bu sistemler Aristo ile başlar Theophrastus ile devam eder ve Herbalist’lerle son bulur Bu ilk dönemde bitkiler dış görünüşlerine göre ağaçlar, çalılar, otlar ve sarılıcılar, tek yıllık, iki yıllık, çok yıllık, petalsiz, çok petalli, tohumlu, tohumsuz vb. gruplandırılmıştır. Bu konuda ilk yazılı eser Aristo’nun öğrencisi Theophrastus (M.Ö. 372-287) tarafından yayınlanmıştır. De Historia Plantarum adlı eserinde yaklaşık 480 kadar bitki çeşidini tanımlamıştır. Bu kişi, bilim aleminde bitki sistematiğinin babası olarak bilinir. M.S. 1. yüzyıl da, Ülkemizde Adana yakınlarında bir yerde yaşayan Dioscorides yaklaşık 600 bitki türü ile bu arada Labiatae (Ballıbabagiller) ve Umbelliferae ((Maydonozgiller) gibi familyaları da tanımlamıştır. Theophrastus 450 kadar kültür bitkisini ot, ağaççık ve ağaç olarak sınıflandırmıştır Daha sonra Adana-Anavarza’lı Dioscorides Materia Medica adlı yapıtında ve Plinius faydalı bitkiler konusunda yazdığı 37 ciltlik yapıtında sistematiğe de yer vermişlerdir. Albertus Magnus (M.S. 1193-1280) diğer önemli bir araştırmacı olup evvelki araştırıcıların yolundan gitmekle birlikte, bitkileri tek ve iki çenekli olmak üzere de sınıflandırmıştır. 1500’lü yıllarda ise Herbalist’ler ortaya çıkmış ve bunlar bitkilerin çok güzel resimlerini çizmişlerdir. Bu devir, tanıtıcı, ayrıntılı güzel bitki resimleri devri olarak hatırlanır. Mekanik (Suni=Yapay) Sistemler (1580-1760) Daha sonra Taksonomi 16. yüzyıla kadar ölü devresini yaşamış, bu yüzyılla başlayan 200 yıllık sürede birçok bitki toplanmış ve eserler yazılmıştır. Bu sistemin önemli araştırıcısı İngiliz John Ray (1623-1705) otsu bitkileri Kriptogamae (bugün Yosun ve Eğreltiler diye bilinen) ve Phanerogamae, Tohumlu Bitkiler diye ayırdıktan sonra, John Ray Historia Plantarum adlı yapıtında ilk kez Monokotiledon ve Dikotiledon terimlerini kullanılmıştır. Çiçekli bitkileri Petalodes (taç yapraklı) ve Apetali (taç yapraksız) olarak ikiye ayırmıştır. Yaklaşık 18.000 türün tanımını (betimini) yapmıştır. Ülkemize ilk bilinçli gelen yabancı botanikçi olan Fransız Tournefort (1656-1708) ilk kez cins adını kullanmış ve modern cins kavramını Botaniğe kazandırmıştır. Bu dönem, bitkinin sadece tanınmasının kolay olması için kasten suni olarak düzenlenen sınıflandırma sistemleriyle karakterize edilir. Bitkiler hakkındaki bilgiler yeterli olmadığından, tüm ağırlığın belirli bir karakterin sayısal özelliklerine verilmesi tamamen yapay bir sınıflandırmaya neden olmuştur. Bu dönemin en önemli ismi, sistematiğe ve diğer doğa bilimlerine büyük katkısı olan İsveçli botanikçi Carolus von Linnaeus’dir (Karl fon Linne okunur) (1707-1778). Tipik bir doğa bilimcisi olan Linne, Zooloji, Jeoloji konusunda da önemli eserler vermiş, çok tanınmış bir doğa bilimcisidir. Bilim hayatının bir periyodunu Hollanda da geçirmiştir. Clifford adlı br zengin Hollandalı Linne’yi ülkesine çağırmış ve uzun yıllar orada çalışmalarını desteklemiştir. Linne Hollandadaki çalışması sonucu Clifford’un Bahçesi adlı kitabı yayınlamıştır. Kendisinden çok öğrencileri yeryüzünün bir çok kesimini dolaşarak bitki toplamış ve bunları kendi sistemlerine göre sınıflandırarak, bugün de kullanılan binomial sisteme göre adlandırmıştır. Linne’nin suni sınıflandırma sisteminin şu anda tarihi değeri dışında bilimsel bir değeri olmamakla birlikte Binominal sistem denen bitkilerin iki isimle adlandırılması halen kullanılan önemli bir prensiptir. John Ray (1627-1705) ve nihayet Carl von Linneaus (1707-1778) bitki ve hayvanların sınıflandırılmalarına yeni kurallar getirdiler. Linneaus 1753’de yayınladığı Species Plantarum (Bitkilerin türleri) adlı eserden sonra bitkiler, 1758’de yayınladığı Systema Natura (Doğa Sistemi) adlı eserinde ise hayvanlar iki isimle (Binominal) adlandırılarak yayınlanmış ve canlılar bu tarihten sonra latince olan iki isimle adlandırılmışlardır. Bu metot günümüzde de devam etmektedir. 1753 yılı bitkilerin iki adla isimlendirilmesinde milad kabul edilir. Bu tarihten önce verilen isimler geçersizdir Linne bitkileri salt eşey organlarının sayısına göre sınıflandırmıştır. Oldukça yapay olan bu sınıflandırmada Linne, bitkileri önce erkek organ sayısına göre 24 sınıfa ve dişi organ sayısına göre de 67 ordoya ayırmıştır. Böylece doğal olarak birbirine hiç benzemeyen bitkiler bir araya gelmiştir. Diandria: Veronica (Scrophulariaceae), Salvia (Lamiaceae) Trandria: Iris (Iridaceae), Bergia (Elatinaceae), ve tüm Poaceae Tetrandria: Mentha (Lamiaceae), Ulmus (Ulmaceae) ve Cornus (Cornaceae). Linne’nin, bitki sistematiği konusunda en önemli eserleri Systema Natura (1735), Genera Plantarum (1737) ve Species Plantarum (1753) dur. Özellikle Species Plantarum (Bitkilerin Türleri) adlı eserin ilk baskısının yayın tarihi olan 1753 binominal sistemin uygulanma başlangıcı olması açısından çok önemlidir. Natural= Doğal sistemler: (1760-1880) Bu devirdeki sistematikçiler artık birkaç karaktere göre sınıflandırma yapılamayacağını anlayarak mümkün olduğunca fazla karakter kullanarak bitkileri doğal olarak sınıflandırmaya başlamışlardır. Michel Adanson (1727-1806) ile başlayan ve nümerik taksonominin babası da sayılan Adanson çok sayıda karakteri kullanıp bugün bilinen çok sayıdaki ordo ve familyaları birbirinden ayırt edebilmiş ve gruplandırmıştır. Bernard De Jussieu (1699-1777) ve yeğeni A.L. De Jussieu (1748-1836), bunlardan sonra De Candolle’ler (1778-1893) dönemi, Robert Brown (1773-1858) kendi sistemlerini kurup, bitkiler bugünkü gruplara çok yakın kategorilere ayırmışlardır. Bu devrin en önemli sistemi iki İngiliz tarafından ortaya atılan ve onların adları ile bilinen Bentham-Hooker sistemidir. Bu sistem bir diğer İngiliz araştırıcı Darvin dönemine rastlamakla beraber Darvin 1859’da Türlerin Kökeni adıl eseri yayınladığında araştırıcılar sistemlerinin yarısına ulaşmışlardı. Bu nedenle sistemlerine evrimi yansıtmadılar. Ancak büyük bir tesadüf eseri sistemleri bugün kullanılan filogenetik sistemlerle büyük benzerlik göstermekle birlikte, çam (Pinus) ve benzer kozalaklı bitkilerin tek ve iki çenekliler arasına konması zamanımız sistemleri ile çelişkilidir. Ancak bu sistem halen Avrupa’nın, İngiltere başta olmak üzere bazı büyük herbaryumları (kurutulmuş bitki örneklerinin saklandığı müzeler) tarafından kullanılmaktadır. 19. yüzyıl başlarında dünyanın her yanından gittikçe daha çok toplanan bitkilerin, Avrupa’nın botanik müzelerinde (botanik bahçe ve herbaryumlarında) birikmesi ve yeni araştırma aletlerinin (örneğin mikroskop) keşfi bitkiler hakkındaki bilgilerimizin artışına yol açtı. Böylece sınıflandırmada sadece belirli bir karaktere dayanmaktansa tüm organlardaki (vegetatif ve floral) benzerliklere dayanmanın daha doğal sistemleri doğurduğu anlaşıldı. Bu dönemin taksonomistlerinden M.Adanson (1727-1806) bütün yapay sistemleri reddetmiş ve aşağı yukarı günümüz ordo ve familyalarının eşdeğeri olan taksonları "Familles des Plantes" adlı eserinde tanımlamıştır. "Lamarkizm" teorisiyle iyi tanıdığımız J.B.A.P.M. de Lamarck (1744-1829) ilk adlandırma anahtarını hazırlayan taksonomisttir. De Jussieu’lar (üç kardeş ve bir oğul) bitkileri bir ve iki çenekli, ovaryum durumu, petalin varlığı veya yokluğu, serbest veya bileşik oluşu gibi özelliklere dayanarak sınıflandırdılar. Birçok bitkiyi bugün bildiğimiz gibi sınıflandırmalarına karşın, birçoğunu da doğal olmayan bir şekilde bir araya getirmişlerdir. Bu devrede daha birçok taksonomist sınıflandırma şekilleri geliştirmişlerdir. Örneğin dede, baba, oğul de Candolle’lar, Bentham ve Hooker gibi. Her taksonomistle, bitki sınıflandırması bir adım daha ilerlemiş ve günümüzdeki bilgilere yaklaşmıştır. Darwin’in The Origin of Species adlı eseri ve Wallace’nin araştırmaları bu dönemi otomatikman kapatmıştır. Çağdaş Filogenetik Sistemler (1880-.............) Bu dönem Darwin’in yayınladığı "Türlerin Kökeni" (Origine of Species) adlı eserin zamanın araştırıcıları tarafından kabul edilmeye başlanması ile ortaya çıkmış ve devrin sistematikçileri Aristo zamanında ortaya atılan ve a zamanlara kadar kabul edilen "Türler başlangıçta ne kadar yaratılmışlarsa şimdi de o kadar çeşit halindedirler" mantığı yerine türlerin zaman içinde evrimleşerek değişebileceği görüşünü kabul edip, sistemlerini bu fikre dayandırarak yapmışlardır.Bu devir sistematikçileri, daha evvelce yapılan taksonomik grupları akraba ilişkilerine göre incelemeye başlamışlar ve "soy ağacı" belirlemeye, bitkileri basitten gelişmişe doğru ve biribirleri ile akrabalık ilişkilerine göre gruplamaya çalışmışlardır. Echler, Engler, Prantl, Rendle, Wettstein, Hutchinson, Soo, Emberger, Tahtajan, Cronquist bu sistemin en önemli isimleridir. Bu araştırıcılar daha evvel saptanan gruplarda çok büyük değişiklikler yapmamakla birlikte, bunları her kategori düzeyinde akrabalık ilişkileri içinde ve sırasında, ilkel sayılanlardan gelişmişe (basitten evrimleşmişe) doğru bir sıra ile incelemişlerdir. Bazı Önemli Taksonomik Prensipler: Sistematikte temel kategori olan türün tarifi çok çeşitli yapılmakla birlikte en çok kabul edilen şudur. Tür "yapısal ve işlevsel özellikleri birbirine benzeyen, aynı dış ve iç kimyasal, fizksel koşullara benzer tepkiler gösteren, doğal koşullarda çiftleşerek verimli yavrular meydana getiren bireyler topluluğu" dur. Darwin’in evrim teorisinin ve daha sonraki araştırmalarının iki önemli sonucu olmuştur. Türler değişmez birimler değil, aksine daima değişen bir devamlılık içindedir. Ortak ataya sahip türler, bir sistematik grupta toplanmalıdır ve gruplar filogenetik (Taksonların kökeni ve evrimi ile ilgili) olarak düzenlenmelidir. Türler, belirli bir tipi gösteren bireyler halinde değil, değişken (çeşitlilik gösteren) populasyonlar halindedir. Böylece bu dönemde, bitkilerin gösterdiği karakterlerin evrimsel gelişimleri öğrenilmeye çalışıldı. Hangi özelliklerin ilkel, hangilerinin gelişmiş olabileceği, (örneğin monokotillik mi ilkeldir yoksa dikotillik mi?) ayrıntılı olarak incelendi ve bazı esaslarda anlaşma sağlandı. Bunlara Paleobotanikteki verilerde eklenerek bitki türlerinin ve daha üst hiyerarşideki (cins, familya vd.) taksonların kökenleri ve akrabalık ilişkileri yani filogenileri saptanmaya çalışıldı. Bu şekilde ortak atadan gelen taksonlar aynı gruplarda toplandı ve filogenetik sistemler ortaya çıktı.Bu devir taksonomistlerin ortaya attıkları sistemler arasında az veya çok farklılıklar vardır. Zamanımızın botanikçileri filogenetik yaklaşımı kabul etmekle birlikte, bugüne kadar dünyada bütün taksonomistlerin fikir birliği içinde olduğu veya çoğunluğun kabul edebileceği bir sistem ortaya konamamıştır.
-
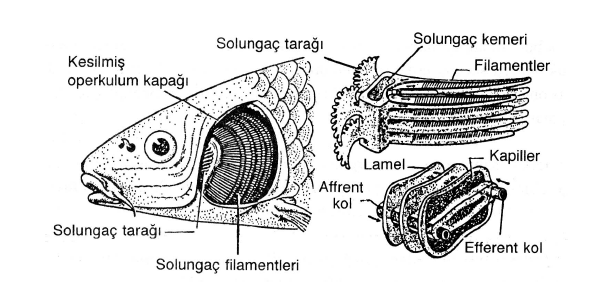
Suda yaşayan hayvanlarda görülen genel solunum sistemi solungaçlardır. Omurgalılardan balıklar, kurbağaların larvaları, omurgasızlardan yumuşakçalar, suda yaşayan bazı solucanlar, eklembacaklılardan karidesler, yengeçler bu yapılara sahiptirler. Bunlar kendiliğinden hareket eden ya da su akımları ile hareket edebilen bir sistem şeklindedir. Omurgalı hayvanlarda solungaçlar akciğerli balıklarda, semenderlerde ve kurbağa larvalarında olduğu gibi vücut dışında olabileceği gibi, bir oda içinde kapalı da olabilirler. Solungaçlar ince epitel çıkıntılarından oluşurlar. Solungaçların üzeri dolaşım sisteminin kılcal damarları ile örülmüştür. Gaz değişimi, solungaç epiteli ve kılcal damarların tek tabakalı yassı eepitelleri arasında difüzyonla gerçekleşir. Suda erimiş oksijen solungaç epitelinden kılcal damarlara geçer. Karbondioksit ise ters yönde hareket eder. Solungacı olan her hayvan, bu organın üzerinde su akımını sağlayan bazı yapılara sahiptir. Örneğin balıklar ağzını açarak bir miktar su alır, sonra ağzını kapatarak ve ağız boşluğunu daraltarak suyun solungaçlar üzerinden geçmesini sağlarlar. Kurbağalar gibi bazı hayvan gruplarında birden fazla solunum görevi yapan organ bulunduğunu biliyormusunuz? Bazı hayvan grupları bu organların bir kaçı ile solunum yapabildikleri gibi, hayat dönemlerinin farklı dönemlerinde değişik solunum organları ile solunum yapabilmektedirler. Örneğin kurbağalar ergin devrelerinde deri ve akciğerlerle solunum yaparlarken, larva devrelerinde solungaç solunumu yaparlar. Başkalaşım sırasında kaybolan solungaçların yerini akciğerler almaktadır.

-
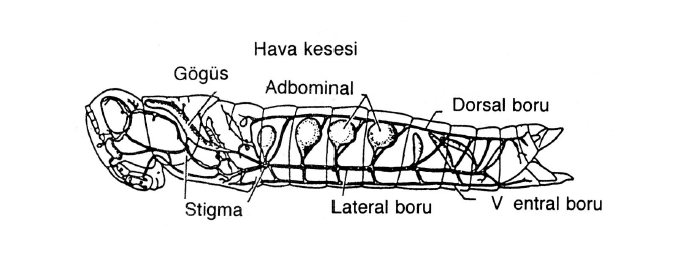
Gelişmiş canlılarda vücut hacmının artması nedeniyle vucudun iç kısımlarında bulunan hücrelerin gaz değişimi ancak özel solunum sistemleriyle gerçekleşebilmektedir. Basit solunum sistemlerinden trakeler bunlardan biridir. Trakeler böceklere özgü, basit borulardan ibaret bir solunum sistemidir. Atmosferik hava, özel borular aracılığı ile vücudun iç hücrelerine iletilerek gaz alış-verişini sağlarlar. Vücudun abdomen kısmında her segmentte bulunan ve stigma adı verilen çift açıklıklardan hava alınıp verilir. Buradan giren hava, içorganlara kadar uzanan dallı hava borularınyla (trakeler) taşınır. Trakeler içi bir sıvı ile dolu ve trakeol adı verilen çok ince borucuklarla sonlanırlar. Oksijen ve karbondioksit değişimi bu sıvı ile dokular arasında gerçekleşir. Böcek hava alışverişini sağlayabilmek için vücudunu kasıp gevşetir. Vücut genişletildiğinde hava trakelere girer, kasılınca dışarı atılır.
-
En ilkel solunum biçimi olan deri solunumunun görüldüğü bazı basit yapılı hayvansal organizmalarda deriden difüzyonla giren oksijen yine difüzyonla diğer doku ve hücrelere iletilir. Basit çok hücreli hayvanlardan yassı solucanlar (Plathelminthes), bazı yuvarlak kurtlar (Nemathelminthes), karasal halkalı solucanlar (Annelida), mikroskobik yapıdaki omurgasız hayvanlar deri solunumunun görüldüğü hayvan gruplarıdır. Omurgalılardan kurbağalarda deri solunum önemli yer tutar. Ayrıca ağzın içini ve yutağı kaplayan zarlar da solunum organı gibi görev yapar. Deri solunum yapan organizmaların derilerinin hep nemli olması gerektiğine dikkat ettiniz mi? Gerçekten de deri solunumu yapan toprak solucanı, salyangozlar ve kurbağa gibi hayvanlarda deri nemli tutulmak zorundadır. Bu nem derideki mukus bezleri ile sağlanmaktadır.
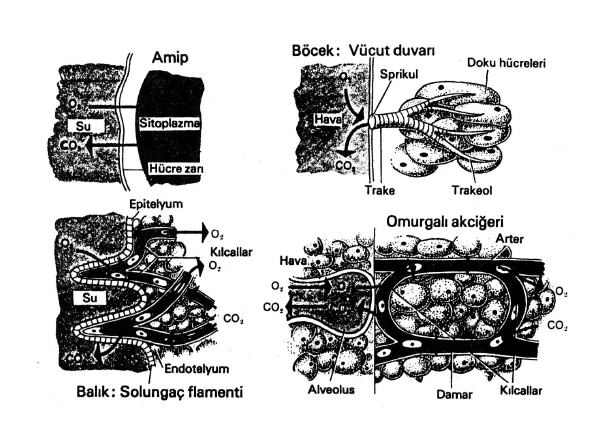
-
Tek hücreli canlılar ile vücut duvarı ince olan süngerler ve sölentereler gibi çok hücreli hayvanlar bu tür gaz değişimlerini özel bir yapıya ihtiyaç duymadan kolaylıkla yapabilirler. Ancak kompleks yapılı olanlar gaz değişimini özel doku ve organlardan oluşan bir sistem aracılığıyla gerçekleştirirler. Çünkü hayvanlarda evrimsel gelişim sonucu her hücrenin dış çevre ile doğrudan doğruya gaz alışverişi yapma olanağı ortadan kalkmıştır. Hayvanlar aleminde solunumda gaz değişimini gerçekleştiren organlar çeşitlilik göstermesine karşılık, bunları dört ana grupta toplamak mümkündür. Dört Genel Tip Solunum Organında Gaz Değişimi a. Hücre Zarı (Bir Hücrelilerde ve Basit çok Hücreli Hayvanlarda) b. Trake (Böceklerde) c. Solungaçlar (Suda Yaşayan Hayvanlarda) d. Akciğer (Karasal Omurgalılarda) Bu değişik sistemlerin ortak yapısal özellikleri ne olabilir, düşününüz? Yanıtınız "gaz alış-verişi için çok geniş yüzeylere sahip olmalarıdır " şeklinde olmalıydı. Ayrıca bu yapıların ince yüzeyli olması, oksijen ve karbondioksidin suda eriyebilmesini sağlayabilmek için nemli tutulmaları ve bol kan almaları zorunludur. Solunumda gaz değişimi iki yerde meydana gelmektedir. Bunlardan birincisi çevre ile solunum organı arasındaki gaz değişimi, ikincisi ise solunum organı ya da taşıma sistemi ile dokular arasındaki gaz değişimidir. Her iki bölgedeki gaz değişiminin esası difüzyon olayına dayanır. Yani değişen gaz, konsantrasyonu çok olduğu yerden, düşük olduğu yere geçer.
-
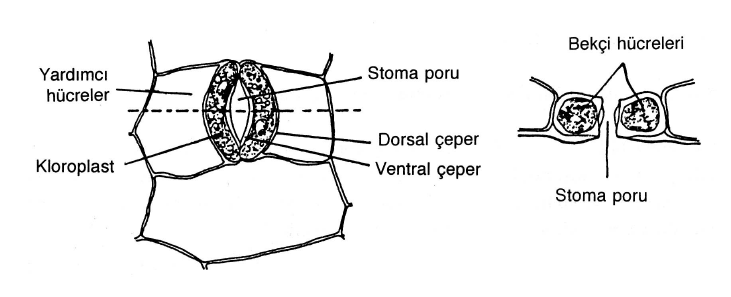
Stomalardan gaz değişiminin sağlanması stomaların açılıp kapanmasıyla gerçekleştirilir. Bu mekanizmanın çalışması hem stoma hücrelerinin hem de çevrelerindeki epidermis ve mezofil hücrelerinin turgor durumuna bağlıdır. Bu hücrelerdeki osmotik konsantrasyonun değişmesi ile oluşan difüzyon farkına bağlı olarak su, ya stoma hücrelerine doğru ya da stoma hücrelerinden komşu epiderma hücrelerine doğru hareket eder. Su komşu epidermis hücrelerinden stoma hücrelerine doğru hareket ettiği zaman stoma hücrelerinin turgor basıncı artar ve stoma porunun daha ince olan dorsal çeperi (epidermis hücreleri ile komşu olan çeper) daha kalın olan ventral çeperden (por tarafındaki çeper) daha çok gerilir. Bu durum daha kalın olan ventral çeperin daha konkav bir şekil almasına ve iki stoma hücresi arasında bir açıklık oluşmasına neden olur. Böylece stoma poru açılır. Aksi durumda, yani stoma hücreleri turgor özelliklerini kaybedince durum tersine döner. Dorsal çeper gerginliğini ventral çeper de konkavlığını kaybeder. Sonunda stoma hücreleri gevşek bir hal alır, aradaki açıklık da kapanmış olur. Buradaki turgor durumunun sağlanması ya da kaybedilmesini sağlayan koşullar ışık, su azlığı ve sıcaklıktır. Su, sıcaklık ve CO2 ve O2 sınırlayıcı birer faktör olmadıkça genel olarak stomalar ışıkta açılır, karanlıkta ise kapanır. Diğer bir söylemle stomalar gündüz açık, gece kapalıdır. Bitkilerin eğer transpirasyonla (terleme) yitirdikleri su miktarı köklerle alınan su miktarından fazla ise bitkide su sıkıntısı başgösterir. Bitkide su sıkıntısının başlaması, stoma hücreleri ile çevrelerindeki mezofil ve epidermis hücreleri arasında bir difüzyon basınç farkı (emme kuvveti) meydana gelmesine neden olur ve bunun sonucu su, stoma hücrelerinden dışarıya yani epidermis ve mezofil hücrelerine doğru hareket eder. Sonuçta stoma hücrelerinin turgoru azalır ve por kapanır. Diğer koşullar sabit olmak üzere belirli bir dereceye kadar sıcaklık artışı stoma porunun açılmasına neden olur. Bitkilerde bu değişimi sadece stomalar mı sağlamaktadır? Yaşlanmış bitki gövedeleri su ve gaz geçişine engel olan mantar dokusu ile örtülü olduğundan, iç kısımdaki canlı hücrelerin hayatsal olaylar sonucu meydana gelen artık gazların ve su buharının dış ortamla alışverişini sağlayan peridermal ve lentisel (kovucuk) adı verilen yapılar meydana gelmiştir.
-
Yüksek bitkilerin organlarının dış yüzleri gerek gaz, gerekse suya karşı çok az geçirgen olan ve hücre arası boşlukları bulunmayan epidermis dokusu ile örtülüdür. Bu nedenle dış ortamla iç ortam arasında gaz alış verişi ve bazı maddelerin dışarıya verilişini sağlamak üzere epidermiste yapılar oluşmuştur. Bu yapılara stoma (gözenek) adı verilmektedir. Bir stoma, aralarında stoma açıklığı (por) denen bir açıklık bırakan ve birbirine bağlı iki stoma hücresi’nden oluşur. Bu yapı havalandırma parenkiması ile birlikte havalandırma sistemini meydana getirir. Havalandırma parenkimasında yer alan hücre arası boşlukları boşlukları stoma altında bulunan ve stoma boşluğu denen geniş bir hücre arası boşluğu ve por yoluyla dış ortama bağlanmıştır. Fotpsentez göreviyle ilgili dokuların üzerindeki epidermiste stoma sayısı fazladır. çiçekte ve su bitkilerinde stomalar ya tamamiyle yok olmuştur ya da sayıları çok azalmıştır. köklerde de stomaya rastlanmaz.
-
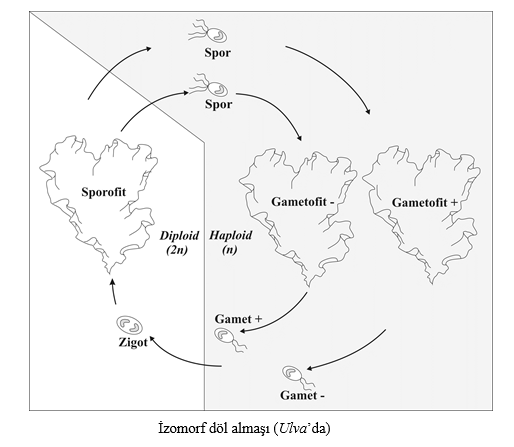
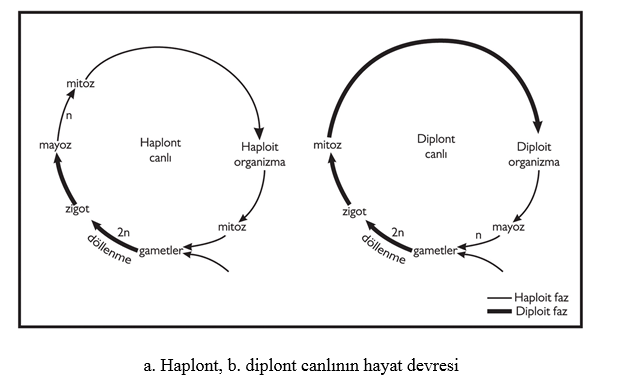
Sınıflandırma başlığı altında da göreceğimiz gibi, algler çok farklı canlı gruplarında yer alan organizmalardır. Aşağıda alglerle ilgili genel bilgilerin yanında sık kullanılan terimlerin tanımlarını da bulacaksınız. Morfolojik Yapılarına Göre Algler Morfolojik olarak farklı alg tiplerini tanımlarken genellikle tek (bir) hücreli, koloni, tallus oluşturan organizmalar gibi terimler kullanılır. Tek hücreli algler, bulundukları ortamda serbest yaşayan bir hücreli bireylerdir. Bulundukları gruba göre değişen sayı ve yapıda kamçı içerebilirler. Bazı tek hücreli algler ise kamçısızdır. Koloni, çok sayıdaki alg bireyinin bir araya gelerek oluşturduğu toplulukları ifade eder. Kolonilerdeki birey sayısı belli cinslerde sabit olabildiği gibi, şekilleri de kendine özgü olabilir. Örneğin kübik, küremsi, tablo şeklinde kolonilerde belirgin şekiller oluşabilir veya bazen de şekilsiz yığınlar halinde koloniler oluşabilir. Alglerle ilgili metinlerde sıkça rastladığımız terimlerden biri de tallus yapısıdır. Fotosentez yapan veya hareket etmeyen tüm organizmaların bitki olarak değerlendirildiği eski dönemlerde kullanılmaya başlayan bu terim kök, gövde, yaprak gibi organ farklılaşması göstermeyen ilkel bitkileri ifade eder. Tallus terimi, bazı mantarlar, karayosunları ve alglerin oluşturduğu yapıları ifade etmek üzere kullanılır. Örneğin yapraksı tallus terimi, yeşil, kırmızı ve kahverengi alglerde görülen yaprağa benzer yapıyı tanımlar. Bazı alg gruplarında görülen parankimatik tallus ise, altta ve üstte epidermis tabakasına benzeyen yassılaşmış, ortada ise parankimatik hücrelere benzeyen tekdüze hücreleri içerir. Ancak talluslu alglerde kesinlikle yüksek bitkilerin yapraklarına benzeyen palizat, sünger parankiması gibi farklılaşmış dokular bulunmaz. Yine sıklıkla kullanılan ipliksi tallus terimi ise ardı ardına dizilen hücrelerden oluşan ve genellikle müsilaj bir kın ya da çeper tarafından çevrilen ince-uzun yapıyı ifade eder. İpliksi bir tallusda yana doğru dallar oluşturulabilir, bu durumda morfolojik yapı dallanmış ipliksi tallus olarak değerlendirilir. Alglerde Üreme ve Yaşam Döngüleri Alglerde eşeyli ve eşeysiz olmak üzere iki tip üreme görülür. Eşeysiz üremede gamet, zigot gibi özelleşmiş hücreler görülmez ve döllenme olmaz. Diğer bütün canlılarda olduğu gibi eşeysiz üreme sonucunda ana bireyin genetik yapı açısından aynısı olan birçok birey meydana gelir. Alglerde eşeysiz üreme, ya spor adı verilen hücrelerin yardımıyla veya tallusun bir parçasının kopup yeni bir birey oluşturmasıyla gerçekleşir. Sporlu eşeysiz üreme, çoğunlukla tek hücreden oluşan eşeysiz sporlarla gerçekleşir. Bu sporlar, genellikle kalın, geçirimsiz çepere sahiptir ve çevre koşulları uygun olduğunda tekrar yeni bireyleri meydana getirirler. Sporlar, ortam koşullarına göre veya ait oldukları alg gruplarına özgü olarak kamçı içerebilir, üretildikleri sporangiumların yapısı ve şekli de türlere göre farklılık gösterebilir. Ana gövdeden bir parçanın kopmasıyla gerçekleşen eşeysiz üreme yöntemi, vejetatif üreme olarak adlandırılır. Bu yöntem de alglerde çok yaygındır. Eşeysiz üreme, hangi yöntemle gerçekleşirse gerçekleşsin, ortamda hızlı yayılmayı ve birey sayısında artışı sağlar. Buna karşın, eşeyli üremeyle genetik çeşitlilik sağlandığı için değişik ortam şartlarına uyum ve neslin devamının sağlanması açısından önemi tartışılmaz Hayat döngüleri açısından değerlendirdiğimizde ise, alglerde haplont, diplont ve diplohaplont olmak üzere üç faklı tip ayırt ederiz: Haplont canlılarda diploid evre sadece zigot oluşumu sırasında görülür. Zigot mayoz bölünme geçirerek yeni bir bireyi oluşturduğu için organizma haplodiddir. Bu organizma mitoz bölünme ile gametlerini verir ve tekrar zigot oluşur, böylece döngü tamamlanır (Şekil 2.1 a). Diplont canlılarda ise, mayoz bölünme gametlerin oluşturulması aşamasında gerçekleşir. Gametler birleşerek zigot oluşur ve zigot da yeni bireyi verir. Diplohaplont canlılarda diploid sporofit ve haploid gametofit olmak üzere iki nesil vardır. Bu iki nesil birbirini izler ve hayat devresi (döl almaşı) tamamlanır. Sporofit nesil üzerinde, genellilke sporangumların içinde, mayoz bölünme ile oluşturulan sporlardan gametofit nesil gelişir. Gametofit nesil olgunlaştığında, üzerindeki genellikle gametangium adı verilen yapılarda üretilen gametlerin birleştiği döllenme olayının sonunda zigot oluşur. Zigottan mitoz bölünme ile yeni bir sporofit nesil meydana gelir. Böylece iki nesilli hayat devresi tamamlanır. Bazı yeşil alg grupları ile kahverengi ve kırmızı alglerde bu tip hayat devresi yaygındır. Daha ilerideki bitki gruplarında da diplohaplont hayat devreleri, çok farklı tipte özelleşmiş organ ve yapılarla sürer. Diplohaplontlar canlılardaki hayat devresinde, eğer haploid gametofit ve diploid sporofit nesiller morfolojik olarak birbirinden ayırt edilemiyorsa, izomorf döl almaşı olarak adlandırılır (Şekil 2.2). Gametofit ve sporofit nesiller morfolojik olarak farklı ise, heteromorf döl almaşı olarak tanımlanır.
-
Bilim insanları uzun yıllar boyunca bilimsel anlamda “bitki” tanımını yapmak üzere tartışmışlar, çeşitli dönemlerde eldeki bilgi birikimine uygun tanımlar ortaya koymuşlardır. İlk yıllarda hareketsiz, yeşil, yapraksı canlılar bitki olarak nitelendirilmiştir. Bu nedenle de fotosentez yapan veya hareketsiz bütün canlılar “Bitkiler Alemi” içinde sınıflandırılmıştır. Algler, mantarlar, küfler, mayalar, cıvık mantarlar, pek çok protozoa üyesi, karayosunları, eğreltiler ve yüksek bitkilerin “bitki” olduğu kabul edilmiştir. Daha sonraki yıllarda, akrabalık ilişkilerini ortaya koymayı hedefleyen sınıflandırma sistemleri geliştirildikten sonra canlıların sınıflandırılmasında pek çok değişiklikler yapılmıştır. Algler, karayosunları, eğreltiler ve tohumlu bitkileri içeren bu anlayışla gelişen sınıflandırma yöntemleri sonucunda vazgeçilmiş, alglerin aslında monofiletik olmayan pek çok grupta değerlendirilmesi gerektiği kabul edilmiştir. Günümüzde alg tanımı, fotosentetik, basit, tek hücreli, koloni oluşturan veya parankimatik talluslu, pek çok farklı taksonomik grupta yer alan canlıları içerir. Alglerin büyük bir çoğunluğu ototrof canlılardır. Bunun yanında heterotrofik beslenme şekilleri de görülebilir. Algler, fotosentez için gerekli olan klorofilin yanında değişik yardımcı pigmentler içerirler. Bu yardımcı pigmentler farklı dalga boylarındaki ışığı adsorblayabildikleri için alg gruplarının sudaki derinliğe bağlı dağılışı da farklılık gösterir. Genellikle sucul ortamlarda gelişen algler, primer üreticiler olmaları nedeniyle yeryüzünde ekolojik açıdan son derece değerlidir. Sucul ortamlarda fotosentez sayesinde oksijen dengesinin korunmasının yanında, algler pek çok omurgasız hayvan ve balık türünün besin kaynağıdır. Ünitemizde incelediğimiz alg gruplarının yeri geldikçe ekonomik özelliklerine de değineceğiz.
-
Pteridophyta (Eğreltiler), çok sayıda fosil formu içeren ve bazı grupları sadece fosil örneklerinden bilinen bir bölümdür. Örneğin, Psilophyton ve Rhynia Devoniendeki kaya fosillerinden bilinen örnekleridir. Vasküler bitkilerin ilkel ve tohum oluşturmayan grubu olarak nitelendirilirler. Pteridophyta’nın ardından Bitkiler Alemi’nin en gelişmiş grubu olan Spermatophyta (Tohumlu Bitkiler) gelir. Eğreltiler, spor oluşturmaları, gametofit neslin ayrı olması, üremede suya bağımlılık gibi ilkel karakterlerin yanında karasal hayata uyumla ilgili önemli adaptasyonlara sahiptir ve kormus yapısı gösterirler. Eğreltilerde bitkinin hayat devresinde baskın olan ve gözümüzle gördüğümüz nesil, sporofit nesildir. Sporofitte görülen organ farklılaşması nedeniyle morfolojik yapıları, yüksek bitkilere çok benzer. Bütün eğreltilerin sporofitinde, ksilem ve floemden oluşan iletim dokusu vardır. Ksilem trakeidlerden ve parankima dokusundan meydana gelmiştir. Floem ise kalburlu borular ve floem parankimasından oluşur. Eğreltilerde farklı tipte yapraklar görülür. Sporofit nesil üzerinde oluşan sporangiumlar, ya trofofil adı verilen özümleme yaprakları veya sporofil adı verilen ve sadece sporları taşımak üzere farklılaşmış yapraklar üzerinde gelişir. Rizom ve köklerinin toprak altında yatay olarak yayılması nedeniyle eğreltiler bulundukları alanda erozyonu önleme ve toprağı tutma açısından öenmli bir işleve sahiptir. Ayrıca halk arasında ilaç, besin kaynağı ve dekoratif amaçlı kullanımları da vardır. Pteridophyta’nın Hayat Devresi Sınıflar arasında farklılıklar görülmekle birlikte, eğrelitlerde hayat devresini genel olarak aşağıdaki şekilde özetlemek mümkündür (Şekil 15). Sporun çimlenmesi ile gametofit nesil gelişmeye başlar. Gruplara göre farklılık göstermekle birlikte gametofit nesil küçük, algsi yapıdadır ve protalyum adını alır. Gametler bu nesil üzerindeki arkegonium ve anteridiumlarda geliştirilir, Erkek gametin yumurta hücresine ulaşması için için suya ihtiyaç duyulur. Döllenmeyle zigot oluşumunun ardından embriyo gelişir. Daha sonra embriyodan kök, gövde, yaprak farklılaşması gösteren sporofit nesli oluşturur. Sporlar, sporangium adı verilen keseciklerin içinde mayoz bölünme ile oluşturulur. Sporangium çeperindeki bazı ölü hücreler asimetrik kalınlaşma gösteren duvarlara sahiptir. Bunlara Annulus hücresi adı verilmektedir. Annulus hücrelerinin bulunduğu gruplarda sporangiumum çeperi bu hücrelerin yer aldığı kısımdan higroskopik hareketlerle açılır ve sporların dağılması sağlanır. Eğreltilerde görülen tipik döl almaşının yanında her iki nesil de vejetatif olarak çoğalabilir. Şimdiye kadar öğrendiklerimizden farklı, bazı özel üreme tipleri görülebilir. Bunlardan biri; gametofitten gamet birleşmesi olmaksızın sporofitin meydana gelmesidir, bu olay apogami olarak adlandırılır. İkincisi ise; sporofittin gamet üretmesi olup, buna da apospori denir. Her iki olay da çeşitli eğrelti gruplarında çok yaygındır. Pteridophyta’nın Sınıflandırılması Eğreltilerin sınıflandırılmasına ilişkin farklı görüşler vardır. Eski yıllarda eğreltilerin tamamı tek bir bölüm “Pteridophyta” içinde sınıflandırmaktaydı. Daha sonra, “Gerçek Eğreltiler” ve yakın grupların ayrı ayrı bölümler içinde değerlendirildiği sistemler düzenlenmiştir. Bu sistemlerde fosil eğreltiler “Psilophyta”, kibritotları “Lycopodiophyta”, atkuyrukları “Equisetophyta, Sphenophyta” ve gerçek eğreltiler “Pteridophyta” olmak üzere genellikle dört divisio yer alır. Günümüzde ise morfolojik karakterlerin yanında moleküler sınıflandırma verileriyle desteklenen akrabalık ilişkilerini de kullanan araştırıcılar ise son yıllarda daha farklı sınıflandırma sistemlerini tercih etmekle birlikte, bu konuda oldukça farklı görüşler vardır. Ünitemizde, yaşayan eğreltileri dört subclassis içinde sınıflandıracağız: Classis 1. Psilotopsida Classis 2. Equisetopsida Classis 3. Marattiopsida Classis 4. Polypodiopsida Classis Psilotopsida Bu sınıfta yer alan ve günümüzde yaşayan cinsler, vasküler bitkilerin en ilkelleri olarak nitelendirilir. Sporofit, köksüzdür, rizoid görülebilir. Gövdedeki indirgenmiş yapraklar çıkıntılar halinde, damarsız veya tek damarlı olabilir, rizomlu bitkilerdir. Sporangiumları büyük, iki hücre kalınlığında çeperli olup annulus hücreleri içermez. Gametofitler ise toprak altında gelişir ve fotosentez yapamadıkları için besinlerini mikoriza oluşturarak sağlar. İki ordoya ayrılan bu sınıfta altı cins yer alır. Ophioglossum, Psilotum ve Tmesipteris en tanınmış cinsleridir. Classis Equisetopsida Sphenophyta adı altında ayrı bir divisio olarak da sınıflandırılan bu grupta günümüzde tropiklerde gelişen yüksek boylu türler dışında alçak boylu, otsu yapıdaki bitkiler bulunur. Paleozoikde yaşayan bazı türlerinin 30 metreye kadar ulaştığı fosil kayıtlarından bilinmektedir. Sporofitlerinde gövde, nodyum (düğüm) ve internodyum (düğüm araları) adı verilen bölümlerden oluşur. Günümüzde yaşayan üyelerin gövdelerinin içinde gençken öz bölgesi bulunur, olgunlaştığında içi boştur. Bu bölgenin etrafında ksilem ve floemin oluşturduğu demetler yer alır. Bu bölümün üyelerinin çoğunda rizom adı verilen toprak altı gövdeleri bulunur. Sporlar, sporofil başakları içinde yer alan sporangiumlarda üretilir. Sporangiumlar annulus içermez, spiral şekilli çeper kalınlaşması gösterir. Olgun sporlar dört adet, kaşık şeklinde elater adı verilen uzun çıkıntılar tarafından sarılmıştır. Elaterlerin açılmasıyla sporlar sporangiumdan atılır ve rüzgarla dağılır. Equisetales tek ordosu, Equisetum ise tek cinsidir. Bu cinsin 30 civarında türü vardır. Equisetaceae familyasının özelliklerini ülkemizde de nemli alanlarda, orman altlarında yaygın olarak bulunan Equisetum örneği ile inceleyelim: Boyları bir metreye kadar ulaşabilen, kısmen kara, kısmen de bataklık bitkileridir. Sürünücü rizomlarından toprak üstü sürgünleri ve gerçek kökleri çıkar. Gövde, oluklu sütun şekli ile karakteristiktir. Nodyum ve internodyumlar çok belirgindir. İletim demetleri düzenli daire şeklinde dizilmiş olup demetlerin içinde de ayrıca boşluklar vardır. Demetlerde floem daha iyi gelişmiştir. Yapraklar iyice indirgenmiş olup brakte halini almıştır, bu yüzden gövde de fotosentez yüzeyini artırmak üzere görev üstlenmiştir. Sporangiyumlar, verimli dalların ucunda sıkı başaklar oluşturacak şekilde dizilmiş olan sporofillerde yerleşmiştir. Her bir sporofilin alt yüzünde, beş ila on adet torba şeklinde sporangiyum bulunur. İzosporik bitkilerdir. Bazı türlerde toprak üstü organlarda dimorfizm görülür: İlkbahar başlangıcında klorofilce fakir ve dallanmayan verimli sürgünler meydana gelir. Bu sürgünler, uçlarında yer alan başaklardaki sporlar döktükten sonra ölürler. Daha sonra, verimsiz ve fazla dallanmış yeşil asimileme sürgünleri oluşur. Classis Marattiopsida Fosilleri Karbonifere dayanan bir gruptur. Kökleri büyük, etlidir, kök, gövde ve yapraklarında müsilaj kanalları bulunur, Rizomları etlidir, çok sayıda sürgün üretir, gövde üzerinde çok büyük tüysü parçalanmış yaprakları bulunur. Sporangiumlarda annulus hücresi bulunmaz, yaprakların altında yerleşmişlerdir ve 1000-7000 spor üretirler. Gametofitleri yeşil olup toprak üzerindedir. Marattiales tek ordosu, Marattiaceae tek familyasıdır ve altı cinsi vardır. Psaronius kömür yataklarında fosilleri bulunan bir cinsidir. Classis Polypodiopsida Sporlu bitkilerin en gelişmiş örneklerini içeren “gerçek eğreltiler”, sporlu bitkiler arasında tohumlu bitkilere en yakın gruptur. Pteridopsida, Filicopsida olarak da isimlendirilmiştir. Sporangiumlarının bir tek epidermal hücreden gelişmesi nedeniyle bu grup “Leptosporangiate” olarak adlandırılır. Bazı üyelerde sporangium toplulukları, indusium adı verilen kalkan şeklinde bir yapı tarafından korunur ve sporangiumlarının etrafında sporların dağılmasını sağlayan annulus hücreleri bulunur. Gerçek eğreltiler, üremelerindeki suya bağımlılık nedeniyle genellikle nemli habitatları tercih eden, oldukça geniş yayılış alanına sahip, toprakta gelişenlerin yanında epifitik türleri de bulunan bitkilerdir.
-
Bryopsida Sınıfı üyeleri morfolojik farklarına göre 3 alt sınıf (subclassis) altında toplanmıştır. Subclassis 1. Sphagnidae Subclassis 2. Andreaeidae Subclassis 3. Bryidae Şimdi bu altsınıfların özelliklerini ve önemli örneklerini inceleyelim. Subclassis. Sphagnidae Bu alt sınıf üyeleri turba yosunları olarak bilinirler. Protonemaları geniş bir tallus yapısındadır. Gametofitin yaprakları midrib içermez, su tutma kapasitesi olan şeffaf ve yeşil renkli iki tip hücreden oluşurlar. Anteridyumlar yaprak koltuklarında, arkegonyum ise gövde ve yan sürgünlerin uç kısımlarında meydana gelir. Bu grubun üyeleri bataklıklarda, sulak alanlarda gelişir. Gametofitin büyümesi yıldan yıla devam eder bu sırada ölen dip kısımlar su içinde birikerek turbalıkları oluşturur. Turba, Kuzey Avrupa ülkelerinde yakacak olarak kullanılır. Su tutma kapasiteleri oldukça fazla olduğu için uzun süre canlı kalması istenen materyallerin paketlenmesinde ve tohumların çimlendirilmesinde de kullanılır. Sphagnales bu subclassisin tek ordosu, Sphagnaceae ise tek familyasıdır. Burada yer alan tek cins olan Sphagnum’un 320 kadar türü vardır. S. compactum Lam. & DC. ve S. palustre L. bu cinsin ülkemizde de yayılış gösteren iki türüdür. Subclassis Andreaeidae Bazı araştırıcılar tarafından Andreaeobrya olarak da adlandırılır. Diğer iki alt sınıfın bazı özelliklerini bir arada taşıyan bir gruptur. Gametofitlerinin yapısı Bryidae üyelerine benzer. Sporogen doku “endotesyum”dan gelişir. Sporofit, kolumella ile sarılıdır. Bu alt sınıf üyelerine has bir özellik, kapsülün 4 kola ayrılarak parçalanmasıdır. Endotesyum: Sporogen dokunun geliştiği tabakanın adıdır. Neuroloma Hedw. ve Andreaea Hedw. burada yer alan iki cinstir. Subclassis Bryidae Bu altsınıf, gerçek karayosunlarını taşır ve Eubrya olarak da adlandırılır. Protonemaları hemen hemen her zaman ipliksi yapıdadır. Yapraklarda midrib vardır. Kapsülü taşıyan sap uzamıştır. Sporogen dokusu endotesyumdan meydana gelir ve kolumella tarafından sarılmaz. Olgun kapsül genellikle birkaç doku tabakası şeklinde farklılaşır. Cinslerin büyük çoğunluğunda, spor çukurları peristom denen bir yapı ile sarılıdır. Bu alt sınıf 650 cins ve 14.000 tür içerir. Fosil türleri de bilinmektedir. Bryidae altsınıfında yer alan ve ülkemizde de yayılış gösteren bazı türler arasında Fissidens bryoides Hedw., Weissia controversa Hedw., Funaria hygrometrica Hedw., Bryum argenteum Hedw., Mnium hornum Hedw. ve Hypnum cupressiforme Hedw. bulunur.
-
Musci, karayosunları olarak da adlandırılan Bryopsida sınıfının 12 000-15 000 civarında türü vardır ve bu sınıf, bölümün en gelişmiş üyelerini içerir. Karayosunlarını diğer Bryophyta üyelerinden ayıran özellikleri aşağıdaki gibi sıralayabiliriz: Karayosunlarının çoğunda gametofit ışınsal simetrilidir. Rizoidleri çok hücreli ve klorofilsizdir. Gametofit daima yapraklı görünümdedir. Mezofil, stoma, petiol vb. gelişmiş kısımlara sahip gerçek yapraklar bulunmaz. Bu yapılara phylloid veya phyllid (yapraksı) adını vermek bilimsel açıdan daha doğru olmakla birlikte, yaprak terimi de yaygın olarak kullanılmaktadır. Yapraklar sarmal olarak dizilir, loplu değildir ve genellikle kosta veya midrib olarak adlandırılan, orta damar diyebileceğimiz bir yapı içerir. Yaprak hücrelerinde bulunan kloroplastlar mercek biçimlidir. Bu grubun arkegonyum ve anteridyumları arasında parafizler bulunur. Sporofit nesildeki spor kapsülünde mnium tip stoma bulunur. Sporları olgunlaştıktan sonra rüzgar yardımıyla boyutlarına göre farklı uzaklıklara yayılabilir. Bryopsida’da Hayat Devresi Bryopsida sınıfının hayat devresinde sporlar çimlenerek, protonemayı (ön çim, ilk iplik) oluşturur, bu yapıdan da gametofit gelişir. Gametofit, yine bitkinin hayat devresinde baskındır, arkegonium ve anteridiumlarda oluşturulan gametlerin döllenmesiyle oluşan zigottan embriyo ve sporofit nesil gelişir. Sporofit neslin, klorofil içermesi ve iyi gelişmiş bir özümleme dokusuna sahip olması nedeniyle gametofite olan bağımlılığı azalmıştır. Ayrıca bu sınıfta kapsül içinde sporların dağılmasını sağlayan özel yapılar bulunmaz. Şimdi, gametofit ve sporofit nesilleri inceleyelim. Bryopsida’da Gametofit Gametofiti verecek olan ipliksi yapıdaki protonema renksiz rizoid ve yapraklı bölümü verecek olan yeşil kloronema olmak üzere iki kısımdan olşur. Protonema, uç kısımlarından hızla büyür. Birkaç hafta içinde 40 cm ye kadar ulaşır ve substrat üzerinde dallanarak yayılır. Bu dönemde yapraklı gametofitin gövde benzeri ekseni oluşturulur. Protonema yapısı, sporun çimlenmesinden başka gametofitin parçalanmasıyla da oluşturulabilir. Doğada bu olaya sık rastlanır. Örneğin; 1 m3 kutup karında 500 den fazla karayosunu fragmenti (parçacık) bulunabilmekte, bunlardan protonema oluşturulmaktadır. Gametofitin gelişmesinin ardından eşeyli üreme organları oluşturulur. Dioik veya monoik türleri içerir. Gametangiumlar genellikle bitkinin uç kısımlarında oluşur. Arkegonium ve anteridium gruplarının arasında parafizler bulunur. Anteridium grupları arasındaki parafizler su alarak şişer ve spermatozoidlerin atılmasına yardım ederler. Bazı türlerin gametofitleri özelleşmiş hücrelerden oluşmuş bir merkezi silindire sahiptir. Bu özelleşmiş hücreler ksilem (hidroidler) ve floemi (leptoidler) andırır. Kalburlu borulara benzeyen leptoidler, şeker taşır, nükleusları yoktur, çok sayıda plazmodezma içerirler. Hidroidler trakeid benzeri hücreler olup ölüdür ve içleri boştur. Su ve çözünmüş mineralleri taşır1ar. Bununla beraber trakeidlerden farklı olarak odunlaşmış sekonder çeperleri yoktur, bu nedenle gametofitin desteklenmesine çok az katkıda bulunurlar. Ayrıca bazı türlerin yapraklarında Mnium tipi stomalara rastlanır. Bryopsida’da Sporofit Döllenmenin ardından zigotun mitoz bölünmeleriyle iğ biçimli embriyo, arkegoniumun içinde gelişmeye başlar. Bu sırada, arkegonium hücreleri bu oluşuma yer açmak üzere bölünür. Üst kısımda bulunan hücrelerin bölünme etkinliğinin sonucunda kaliptra adı verilen koruyucu bir örtü oluşur. Olgunlaşmış bir sporofit, ayak, sap ve kapsül kısımlarını içerir (Şekil 3.12). Ayak, arkegonium haznesinin tabanına kadar uzanır, gametofite tutunur ve ondan su, mineral madde ve besinleri emer. İnce bir yapı olan sap kısmı ise, kapsülü taşır ve bazı türlerde 15 cm’ye kadar uzayabilir. Kapsül ve sapın birleştiği yerdeki şişkin geçiş bölgesinde minium tip stomalar bulunur. Kapsül, içinde sporların gelişeceği yapıdır ve sporları korumaya, beslemeye ve dağıtmaya yardımcı olan kısımlara sahiptir. Kapsül üzerinde bazen kaliptra adı verilen doku bulunabilir. Kapsülün en içindeki hücreler, sporlara su ve besin sağlayan kolumella dokusunu oluşturur. Dıştaki ceket tabakası ve kolumella arasında kalan sporogen hücreler ise mayoz bölünme ile sporları meydana getirirler. Bir kapsülde 50 milyon kadar spor oluşturulabilir. Sporlar olgunlaştığında kaliptra düşer, böylece operkulum kısmı açıkta kalır. Operkulum, kapsülün kurumasıyla düşer. Bu kapağın hemen altı peristom dişleri adını alan higroskopik dişlerle çevrelenmiştir. Sporlar buradan dökülür. Bazı karayosunlarında ise peristom koni şeklindedir ve sporların çıkabileceği porlar içerir. Operkulum: Spor kapsülünün kapak kısmı. Peristom: Spor kapsülünün ağız kısmı.